

- 光合成系の環境応答機構の解明
- 細胞内代謝の環境応答機構の解明
- シアノバクテリアの代謝改変と燃料生産
- 朋子グループの研究内容についてはこちらをご覧ください。

光合成系の環境応答機構の解明
光合成は光エネルギーを用いて二酸化炭素を固定し、糖を作り出す反応ですが、光強度、温度、栄養条件など環境条件の変化の影響を強く受けます。光合成生物はこれらの環境変化に対して、自らの体を作りかえることで光合成反応を最適化すると共に、環境ストレスによるダメージを回避しています。私達は、光合成生物が環境変動をどのように感じ取っているのか、細胞内でどのようなシグナル伝達が行われているのか、その結果として遺伝子発現やタンパク質の活性がどのように調節されて光合成系の環境応答が実現されるのか、分子レベルでのメカニズム解明を目指しています。主に、シアノバクテリアSynechocystis sp. PCC 6803の転写制御メカニズムに着目した研究を行っており、これまでに得られた主な成果は以下のとおりです。
①DNAマイクロアレイを用いた網羅的解析を行い、光強度や光合成活性の変動によって遺伝子発現がどのような影響を受けるかを明らかにした。
 Hihara et al. (2001) Plant Cell 13:793-806
Hihara et al. (2001) Plant Cell 13:793-806
Hihara et al. (2003) J Bacteriol 185:1719-1725
Aurora et al. (2007) OMICS 11:166-185
Muramatsu and Hihara (2012) J Plant Res 125: 11-39
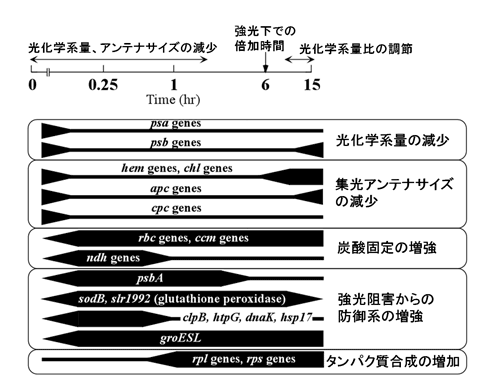
シアノバクテリアSynechocystis sp. PCC 6803を弱光から強光へ移した後に観察される遺伝子発現変動と順化応答のタイムコース
強光条件下においては、遺伝子発現変動とその後に観察される細胞レベルでの応答が密接に関連している。これらの強光応答性を示す遺伝子群の多くは光強度の変化そのものではなく、光強度変化の結果、光合成電子伝達活性が変化することに応答していることが明らかになった。
②転写因子RpaBのDNA結合活性が、光合成電子伝達鎖のレドックス状態に依存して変動することで、光合成関連遺伝子の発現制御が行われていることを明らかにした。
 Muramatsu and Hihara (2003) Planta 216:446-453
Muramatsu and Hihara (2003) Planta 216:446-453
Muramatsu and Hihara (2006) Plant Cell Physiol 47:878-890
Muramatsu and Hihara (2007) J Bacteriol 189:2750-2758
Seino et al. (2009) J Bacteriol 191:1581-1586
Muramatsu et al. (2009) Microbiology 155:989-996
Takahashi et al. (2010) J Bacteriol 192:4031-4036
Kadowaki et al. (2016) Plant Cell Physiol 57:813-823
Riediger et al. (2019) iScience 15: 316-331
Kato et al. (2022) Plant Cell Physiol 63: 1078-1090
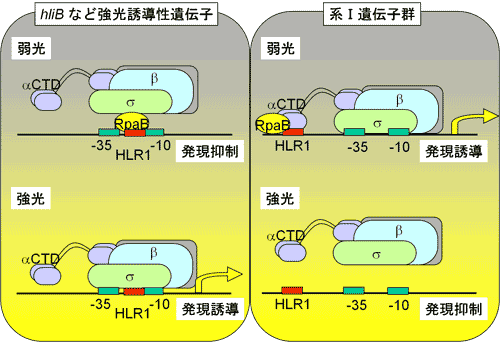
転写因子RpaBによる強光応答性遺伝子の発現制御機構
RpaBはhliBなどの強光誘導性遺伝子の、弱光下での発現抑制に関わる転写因子として報告されていたが、当研究室ではRpaBが光化学系Ⅰ遺伝子(強光下で発現抑制される)の、弱光下での発現誘導に関わることを新たに見い出した。どのようにして同一の転写因子が、ある遺伝子については発現抑制、別の遺伝子については発現誘導に働くことができるのか?そのカギはRpaBが結合するHLR1配列の位置にある。強光誘導性遺伝子の場合、HLR1配列はコアプロモーター領域内に存在し、この領域へのRpaBの結合は、RNAポリメラーゼのアクセスに阻害的に働くと考えられる。一方、系Ⅰ遺伝子の場合はコアプロモーター領域より上流にHLR1配列が存在する。この場合、RpaBの結合はRNAポリメラーゼのアクセスに促進的に働くと考えられる。強光照射により、RpaBはHLR1領域から解離し、結果として強光誘導性遺伝子の発現誘導、系Ⅰ遺伝子の発現抑制が観察される。
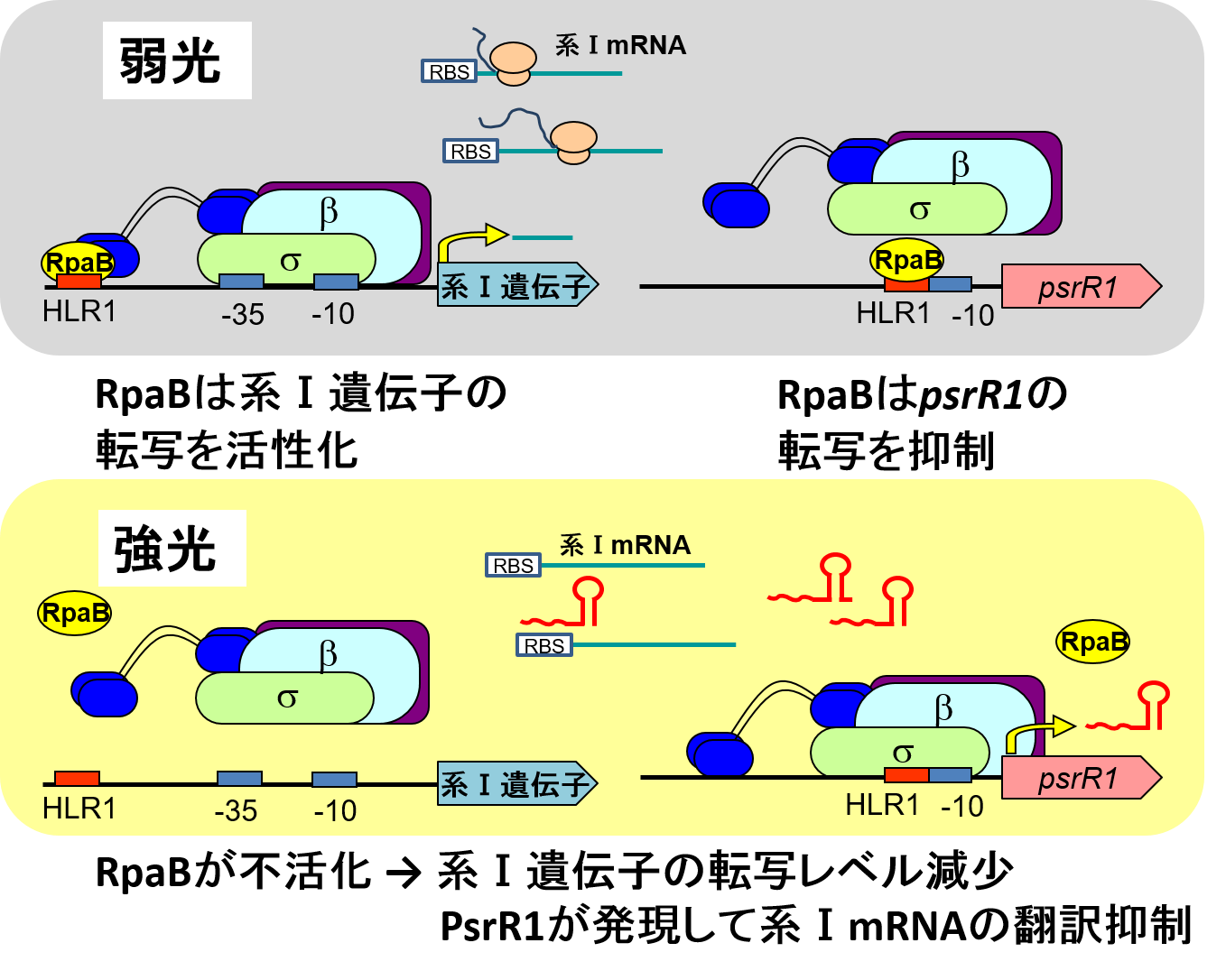
転写因子RpaBと低分子RNA PsrR1による光化学系Ⅰ遺伝子の二重の発現抑制
低分子RNA PsrR1は強光下で誘導され、光化学系Ⅰ転写産物と相互作用して翻訳抑制に働くことが、ドイツFreiburg大学のHess教授・Wilde教授のグループにより示されている(Georg et a. 2014 Plant Cell)。psrR1遺伝子の-10領域にオーバーラップしてHLR1配列が存在し、RpaBが弱光下で転写抑制に働くことが示唆されたことから、Hess教授との共同研究を開始した。その結果、RpaBは上で述べた系Ⅰ遺伝子の直接的制御(弱光で系Ⅰ遺伝子の転写活性化に働き、強光下では不活化して系Ⅰ遺伝子の転写レベルが減少)に加え、PsrR1の転写を弱光下で抑制・強光下では脱抑制することにより、PsrR1による系Ⅰ転写産物の翻訳抑制という間接的制御にも関わることが明らかになった。このRpaBとPsrR1の関係性は"type
4 feed-forward loop”という調節サーキットにあたり、強光下での迅速かつ厳密な系Ⅰ遺伝子の発現抑制を可能にしている。
Hess教授とは、Synechocystis sp. PCC 6803ゲノム上の転写開始点マッピングデータに基づき、転写開始点近傍にHLR1配列を持つ遺伝子をRpaB標的候補としてリストし、そのうちのいくつかについて個別検証を行う共同研究も行った。その結果、RpaBが光合成タンパク質複合体関連遺伝子のみならず、循環的電子伝達に関わるpgrR、光呼吸に関わるgcvP、系Ⅱの修復に関わるプロテアーゼをコードするftsH2など、より広範な光合成関連遺伝子の発現制御に関わることや、窒素代謝のマスター転写因子NtcAや鉄代謝のマスター転写因子FurAなどとともに複雑な制御ネットワークを構成していることが明らかになった。
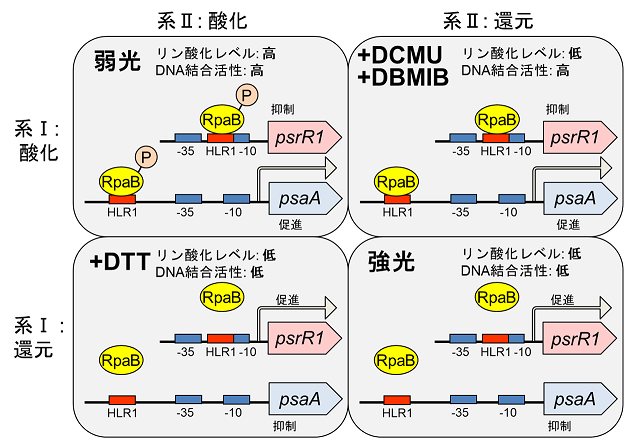
光合成電子伝達鎖のレドックス状態に依存した転写因子RpaBの活性制御
光合成電子伝達鎖のレドックス状態を人為的に変化させ、RpaBのリン酸化レベルとDNA結合活性への影響を調べることで、光化学系Ⅱと系Ⅰのレドックス状態がそれぞれ独立に検知されて、RpaBの活性制御に関わっていることを明らかにした。系Ⅱと系Ⅰの還元は、それぞれ独立に脱リン酸化シグナルとして働き得るが、系Ⅰが酸化的な条件では、リン酸化状態によらず
DNA結合活性は高く維持される。つまりRpaBのDNA結合活性の低下には、系Ⅰの還元が必須であることを明らかにした。
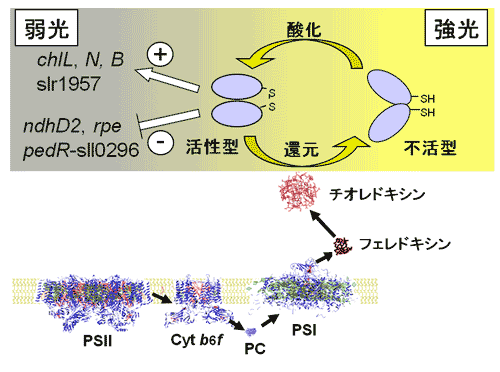
③光合成電子伝達活性に依存した転写制御に働く転写因子PedRを同定し、この転写因子がレドックス制御タンパク質チオレドキシンと相互作用することを明らかにした。さらに、チオレドキシンと相互作用し得る転写因子を複数同定した。
 Nakamura and Hihara (2006) J Biol Chem 281:36758-36766
Nakamura and Hihara (2006) J Biol Chem 281:36758-36766
Horiuchi et al. (2010) Biochem J 431:135-140
Kadowaki et al. (2015) PLoS One 10: e0119107
Kujirai et al. (2018) Sci Rep 8: 6666
転写因子PedRによる強光応答性遺伝子の制御機構
弱光条件下においては、PedRは活性型であり、その標的遺伝子の発現を正または負に制御しているが、強光照射により光合成電子伝達活性が高まると、チオレドキシン経由で還元力を受け取り、構造変化を起こして不活型になると考えられる。
PedRはその後は強光下でも次第に活性型に戻っていき、その標的遺伝子の発現レベルも弱光下に近い状態に戻っていく。
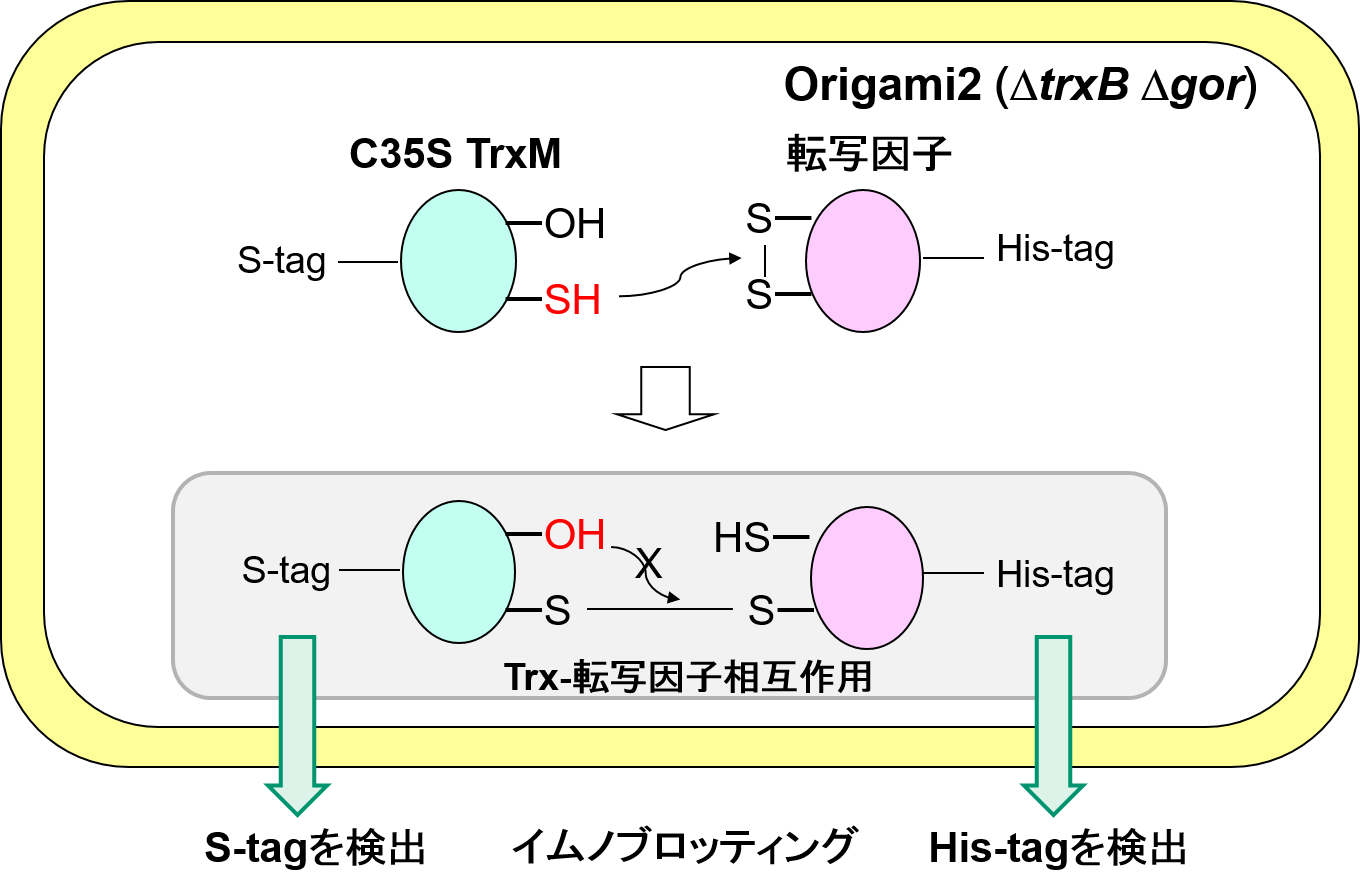
大腸菌共発現系を用いたチオレドキシン(Trx)と転写因子の相互作用の検出
Trxと標的タンパク質の間の相互作用は通常は一過的であるが、Trxの活性部位の二つのシステインのうち一つをセリンに置換する(C35S)ことで、チオール・ジスルフィド変換反応を途中で止めて両者の複合体を検出することができる。我々は、Trxと相互作用する転写因子を同定するため、C35S
TrxMをSタグ付き、および相互作用を調べたい転写因子をHisタグ付きで、大腸菌内で共発現させ、両者が複合体形成するかどうかを、SタグおよびHisタグを検出するイムノブロット解析により調べた。その結果、システイン残基の保存性が高いSynechocystis sp. PCC 6803の転写因子39種のうち、5種を新たなTrx標的候補として同定し、現在その個別解析を進めている。
細胞内代謝の環境応答機構の解明
シアノバクテリアの細胞内では、炭素や窒素等の栄養分の代謝が相互に密接に関連しながら行われています。これらも光合成反応と同様、環境条件の変化の影響を強く受け、栄養分の細胞内への取り込みや代謝経路の活性が様々なレベルで調節を受けていることが分かっています。私達の研究室では、細胞内代謝の調節に関する以下のような研究も行っています。
①光合成と糖異化の両方が活発に行われる光混合栄養条件下では、どのような代謝制御が行われているのか、光混合栄養条件下で致死となるPmgA、Hik31、cyAbrB2などの調節因子の欠損株について遺伝子発現解析や代謝解析を行うことにより、これらの調節因子が担う役割の解明を目指している。
 Kahlon et al. (2006) Microbiology 152:647-655
Kahlon et al. (2006) Microbiology 152:647-655
Takahashi et al. (2008) J Exp Bot 59:3009-3018
Haimovich-Dayan et al. (2011) Environ Microbiol 13:1767-1777
Kaniya et al. (2013) Plant Physiol 162:1153-1163
Nishijima et al. (2015) Photosynth Res 126:465-475
光混合栄養条件下で出現するpmgA破壊株の疑似復帰変異株
pmgA変異株をグルコース添加培地上で光照射して生育させると(光混合栄養条件)、多くの細胞は白化して死滅するが、その中から緑色の疑似復帰変異体のコロニーが現われる。pmgA変異株がなぜ光混合栄養条件下で致死になるのかを明らかにするため、これらの疑似復帰変異体について変異遺伝子を同定したところ、その多くがCO2取り込みに関与するNADPHデヒドロゲナーゼ(NDH-1)複合体のサブユニットに変異を持つことが分かった。
②cyAbrB2転写因子が、炭素・窒素代謝の制御に重要な役割を果たすことを見出し、その機能解析を進めている。
Ishii and Hihara (2008) Plant Physiol 148:660-670
 Lieman-Hurwitz et al. (2009) Environ Microbiol 11:927-936
Lieman-Hurwitz et al. (2009) Environ Microbiol 11:927-936
Yamauchi et al. (2011) J Bacteriol 193:3702-3709
Kaniya et al. (2013) Plant Physiol 162:1153-1163
Hanai et al. (2014) Life 4:770-787
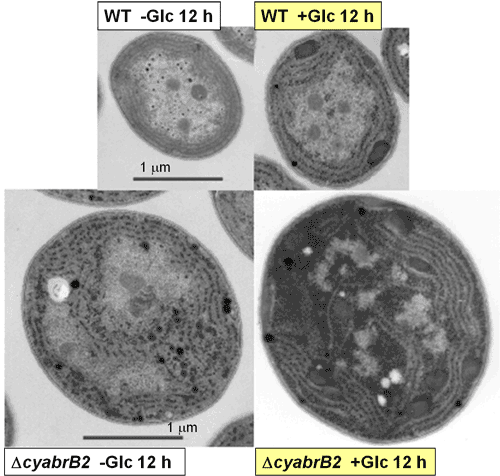
野生株(WT)とcyAbrB2転写因子欠損株(ΔcyabrB2)の電子顕微鏡写真
野生株に比べ、cyAbrB2転写因子欠損株では細胞体積が約5倍となっている。また、光独立栄養条件下(-Glc)において、野生株には見られないグリコーゲン顆粒がcyAbrB2欠損株では顕著に見られる(同心円状に存在するチラコイド膜に沿って蓄積している黒い粒々)。光混合栄養条件下(+Glc)では野生株でもチラコイド膜間にグリコーゲン粒の蓄積が見られる一方、cyAbrB2欠損株ではグリコーゲンがぎっしりと蓄積した結果、細胞全体が黒っぽく見えている。この光混合栄養条件下では、cyAbrB2欠損株は厳しい増殖阻害を受ける。
③シアノバクテリアにおけるLexA転写因子の機能解析を行い、脂肪酸生合成や塩ストレス応答に関わる遺伝子群の発現制御に関わることを見出した。
Kizawa et al. (2016) Front Microbiol 7:193
Kizawa et al. (2017) Plant J 92:189-198
Takashima et al. (2020) Sci Rep 10:17393
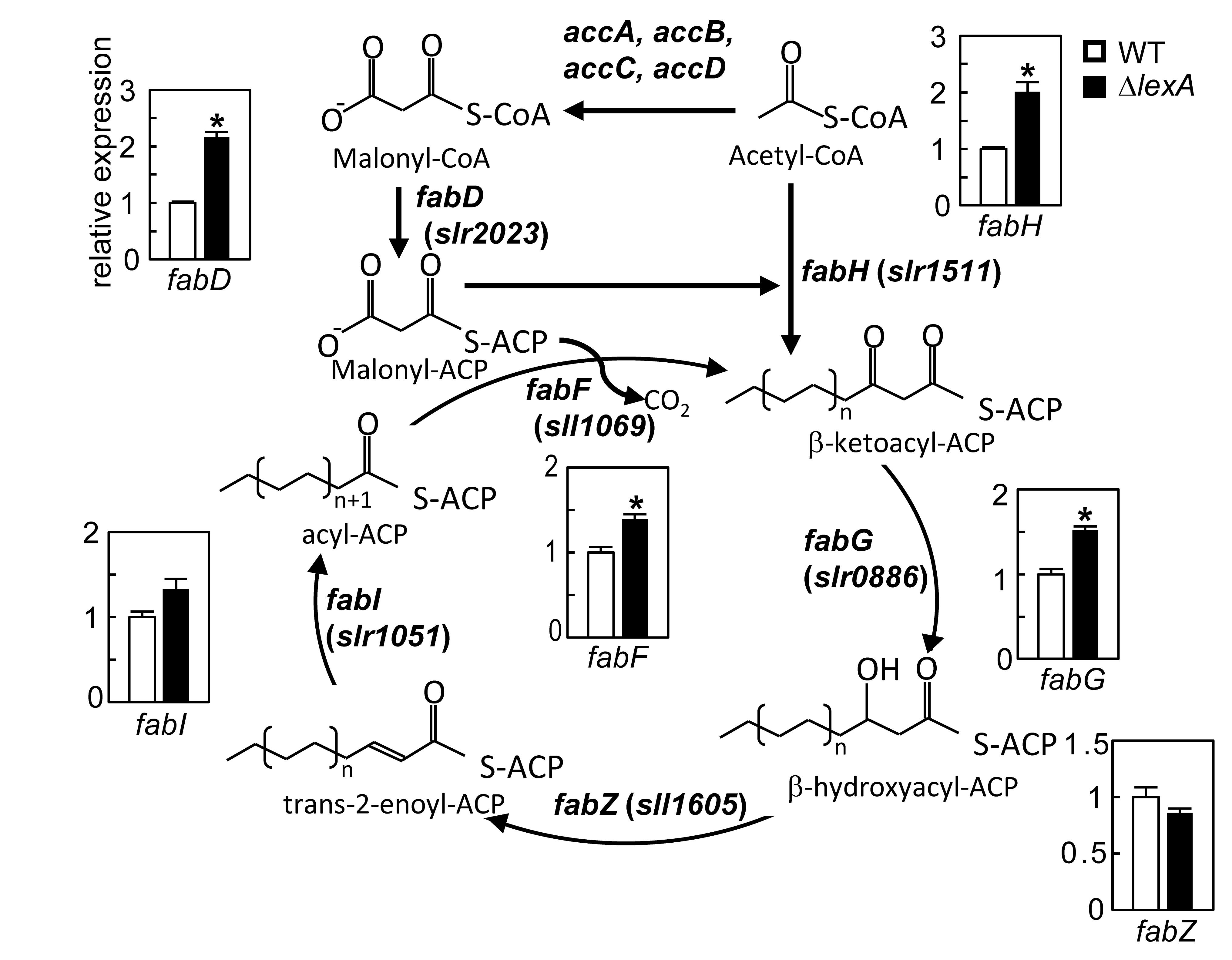
脂肪酸生合成経路の各酵素遺伝子の発現レベルにlexA破壊が及ぼす影響
脂肪酸生合成経路を構成するfab遺伝子群の上流域に結合するタンパク質を単離したところ、LexA転写因子が同定された。欠損株を用いた解析から、LexAは通常条件下で、脂肪酸生合成の開始過程
(fabD、fabH、fabF) および伸長過程の最初の還元反応 (fabG)に働く遺伝子の発現を抑制することが明らかになった。
LexAは一般的には、DNA損傷時のSOS応答に働くことが知られているが、シアノバクテリアSynechocystis sp. PCC 6803では、SOS応答への関わりは報告されておらず、RNA-seq解析を行うと、脂肪酸生合成の他には、pili繊毛の形成、双方向ヒドロゲナーゼの発現、適合溶質グルコシルグリセロールの蓄積等、様々な細胞機能に関わることが明らかになった。
塩ストレス応答に関しては、グルコシルグリセロールの合成と取り込みに関わる4つのオペロンをはじめとする、多くの塩ストレス誘導性遺伝子のリプレッサーとして働くものの、これらの遺伝子の塩ストレス下での誘導はLexAに依存しないこと、一部のLexA分子において173番セリンが塩ストレス下で脱リン酸化されることを見出した。これらの結果から、従属栄養細菌のLexAが自己切断による不活化によりSOS応答を誘導するのとは異なり、シアノバクテリアのLexAは環境条件の変動に対して、リン酸化状態を部分的に変化させることにより、遺伝子発現の微調整に働く可能性が示された。
シアノバクテリアの代謝改変と燃料生産
JST「さきがけ研究」に採択されたことをきっかけに、これまでに行ってきた基礎研究の成果を生かして細胞内代謝を調節し、シアノバクテリアの遊離脂肪酸生産能を高める応用研究を開始しました。その研究の過程でシアノバクテリアの新規中性脂質を同定し、現在、新たな基礎研究を展開しているところです。
①細胞内にグリコーゲンを高蓄積するcyAbrB2欠損株が、燃料生産ホストとして有用であることを示した。
Kawahara et al. (2016) J Biotechnol 220:1-11
Kodama et al. (2018) Biotechnol Bioeng 115: 2974-2985
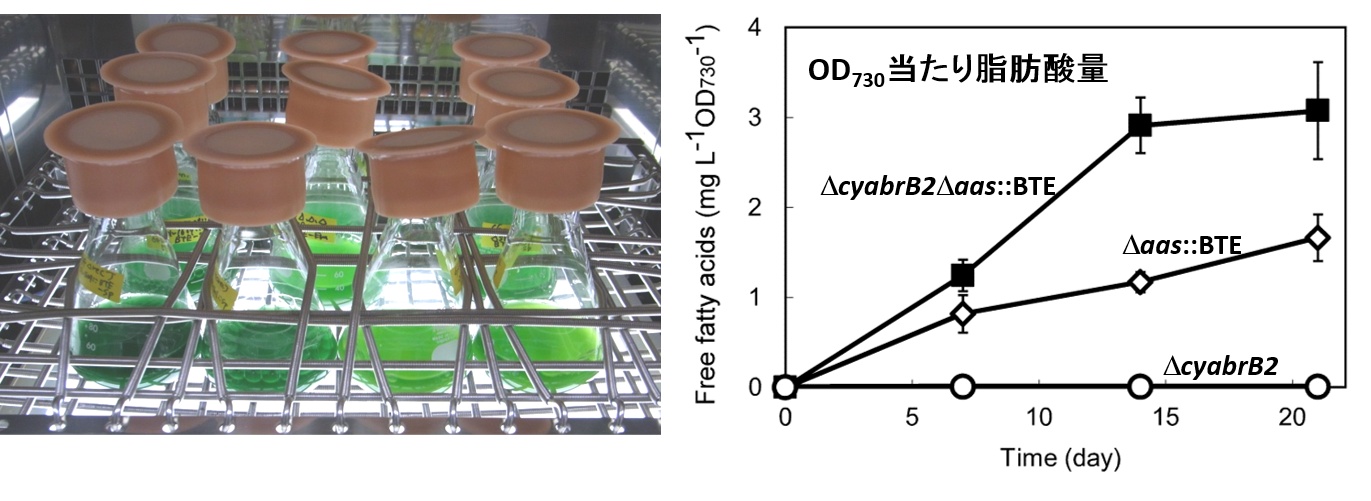
脂肪酸放出代謝改変を施したSynechocystis sp. PCC 6803株の培養
脂肪酸放出代謝改変を施した野生株 (Δaas::BTE)、cyAbrB2欠損株 (ΔcyabrB2Δaas::BTE)、および代謝改変を施していないcyAbrB2欠損株 (ΔcyabrB2)を3週間培養し、培地中への脂肪酸放出量を、細胞濁度当たりの値で示した。cyAbrB2欠損株では、野生株と比べて倍程度の脂肪酸放出が観察された。
②シアノバクテリアが主に蓄積する中性脂質はトリアシルグリセロールではなく、脂肪酸1分子をエステル結合したアシル化プラストキノールであることを見出した。
Tanaka et al. (2020) Plant Cell Physiol 61:1537-1547
Ishikawa et al. (2023) PNAS Nexus 2: pgad092