
Fe-Sクラスターは、in vitro では Fe2+/3+ と S2- から簡単に再構成することができるため、従来、Fe-Sクラスターは非酵素的(化学的、自発的)に生じるものと考えられていました。しかし、今から20年くらい前に私は、葉緑体の中にATP依存的な生合成活性があることに気がつきました。その実体の解明には時間がかかりましたが、大腸菌を用いた遺伝生化学的な解析によって、2種類の生合成系を同定することができました。一つはiscオペロン(iscRSUA-hscBA-fdx-iscX)にコードされるISC(iron-sulfur cluster)マシナリー、もうひとつは、sufABCDSEオペロンにコードされるSUF(sulfur)マシナリーです。すなわち、大腸菌でこれら二つのオペロンのうち片方でも過剰に発現させると、細胞内のクラスター合成活性が高まり、逆にオペロンを破壊すると低下します。二つのオペロンの二重破壊は合成致死となり、もはや生きられなくなります。大腸菌ではISCとSUFの二つの経路が独立して機能していますが、これはむしろ例外的で、他の生き物の多くはどちらか一方だけを持っています。ISCマシナリーはプロテオバクテリアから真核生物のミトコンドリアに見られますが、SUFの方はより広く、真正細菌全般に加えて古細菌や植物の葉緑体にも分布しています。また、一部の嫌気的な生き物には第三の生合成系、NIFマシナリーがあることも明らかにしました。
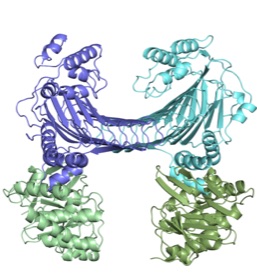
ISCとSUFマシナリーは、どちらも多くの成分から構成される複雑な生合成系です。マシナリーを比較すると、一部の成分には類似性が見られますが、残りの大部分は全く異質なので、反応機構もかなり異なるだろうと予想しています。では、Fe-Sクラスター形成という一見単純な化学反応に、どうしてこれだけ大掛かりな装置が必要なのでしょうか? 逆に言えば、これらのマシナリーの中で、どのようにしてFe-Sクラスターが作られているのでしょうか? この反応機構の解明が現在の最大の課題です。そのためには、マシナリーの個々の成分がどのような性質を持つタンパク質で、どのような役割を演じているのか? また、複数の成分がどのように調和(相互作用)してマシナリーとしての機能を発揮するのか? 分子、原子のレベルで詳しく調べる必要があります。例えば、私たちはこれまで、ISCマシナリーの成分としてFdx、IscX、IscU、SUFマシナリーではSufC、SufD二量体、SufC2SufD2複合体の立体構造をX線結晶解析によって解き明かしましたが、反応機構に関してはまだまだ分からないことだらけです。
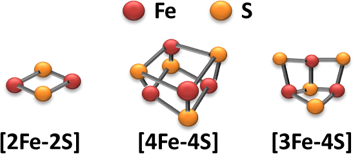
鉄硫黄クラスターと鉄硫黄タンパク質
鉄硫黄(Fe-S)クラスターは、一般に [2Fe-2S] または [3Fe-4S]、[4Fe-4S] の形でタンパク質内部のシステインの硫黄原子に配位結合しています。これらのクラスターをコファクターとして持つタンパク質を総じてFe-Sタンパク質と呼びますが、そのメンバーは細菌から高等動植物まで、150を越える多彩な種類が知られています。生化学の教科書に載っているミトコンドリアの呼吸鎖電子伝達複合体I、II、IIIや、葉緑体のフェレドキシン、光化学系I複合体、シトクロムb6f複合体などは、Fe-Sタンパク質のほんの一部です。これら多くのFe-Sタンパク質は酸化還元(電子伝達)反応に関与していますが、中にはクエン酸回路のアコニターゼのようにLewis酸としての触媒機能を持つものや、ラジカル反応に関与するもの(radical SAM 酵素群)、さらにはFe-Sクラスターをセンサーとして用いる発現制御タンパク質もあります。つまり、Fe-Sタンパク質はエネルギー代謝(光合成、呼吸)や無機物(硫黄、窒素)同化系、ヌクレオチド代謝、アミノ酸代謝、ヘムやクロロフィル、ステロイドホルモン、NADHの合成、DNA修復、環境センサーなど様々な細胞機能においてたいへん重要な役割を果たしています。そういったFe-Sタンパク質の機能を支えているのがFe-Sクラスターの生合成系であり、私たちは、その生合成機構の解明を目的として取り組んでいます。
鉄硫黄クラスター生合成系の遺伝学的・生化学的解析
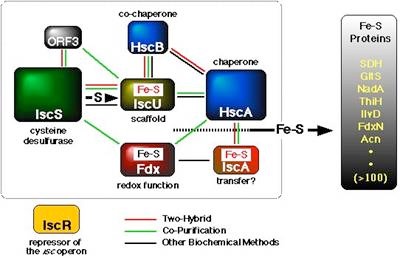
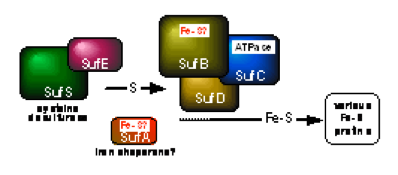
鉄硫黄クラスター生合成マシナリー
究極の目標は、ISCとSUFマシナリーのそれぞれの特性と共通する基本原理を解き明かして、Fe-Sクラスターの生合成機構を詳細に理解することです。そのためには遺伝学、生化学、分光学、構造生物学的な解析を有機的に組み合わせて推進する必要があります。ただし、この生合成反応は、単純な酵素反応とは異質のものです。これまでの知見に基づいて簡単に類推できるようなものではありません。創意工夫と試行錯誤を繰り返しながら道を拓いていくつもりです。
1. 遺伝学的な実験では大腸菌の関連遺伝子群を自在に操り、さまざまな変異の導入によって生じる表現型の解析と、機能不全になった場合にはそれを回復するような二次的サプレッサー変異のスクリーニングを予定しています。言うまでもありませんが、Fe-Sクラスター合成系は大腸菌から高等動植物にいたるまで保存されていますので、基本的なメカニズムの解析には大腸菌が最も使いやすい材料となります。一方、枯草菌やシアノバクテリアやなどにもSUFマシナリーがあり、大腸菌のものとは少し変わっています。朝井先生にご協力頂いて、こういった生き物にも研究を展開できればと考えています。
2. 生化学的な実験ではマシナリーの個々の成分、あるいは複合体を大腸菌で過剰に発現させて精製し、それらの性質を調べることになります。立体構造の解明を目指して結晶化にもチャレンジします。また、構造情報に基づいて部位特異的変異を導入し、その変異タンパク質についても同様に調べていきます。一例を示すと、私たちは最近、IscUの結晶構造に基づいて種々の変異を導入し、それら変異IscUタンパク質の生化学的な性質を調べたところ、結晶構造からは思いもつかない柔軟性を持つということが分かってきました。
研究活動では、学生が時間をかけて自分で勉強し、実験し、その結果について考えることが重要です。主体的に深く掘り下げて考えるという経験は、いずれ自分の力で道を切り拓いて行くための大切な訓練になります。また、学生がアイデアを発揮してこそ、研究は個性的な面白いものになります。私は学生の自主性を尊重しますが、これは放任とは全く異なっており、日常的な討論が不可欠です。教官と学生、学生と学生の間での自由闊達な議論を奨励します。
ポリシー
“もっと詳しく”知りたい方はこちら「詳しい研究内容」へ
もくじ
1.はじめに
2.葉緑体におけるATP依存的なFe-Sクラスター形成活性
3.大腸菌ISC(iron-sulfur cluster)マシナリーの同定
4.新たなFe-Sクラスター合成系:SUF(sulfur)マシナリーの同定
5.多様なFe-Sクラスター合成系の特性と微生物の生存戦略
6.ISCマシナリーの構造と機能
7.SUFマシナリーの構造と機能
8.研究の展望と意義
さらに詳しく
Copyright © 1999- Laboratory of Regulatory Network of Cellular Molecules, Saitama University
