環境応答研究室(西山研究室)
埼玉大学理学部分子生物学科 /
大学院理工学研究科分子生物学コース
JP / EN
RESEARCH
研究テーマ
環境応答研究室(西山研究室)
埼玉大学理学部分子生物学科 /
大学院理工学研究科分子生物学コース
RESEARCH
研究テーマ
光合成生物は、刻々と変動する環境の中で生命活動を維持するために、様々な適応手段を発達させてきました。具体的には、光の強弱や温度の上下といった環境の変化に対応して、光合成機能を最適化するだけでなく、環境変化によって生じるストレスから身を守るための機構も備えています。これらの適応は、環境の変化を感知し、その情報を細胞内に伝達することで、特定の遺伝子の発現を転写、翻訳、および翻訳後レベルで精密に制御することによって可能になります。
当研究室では、シアノバクテリアや真核藻類、植物といった光合成生物の環境応答メカニズムを、遺伝子やタンパク質のレベルで解明することを目指しています。さらに、これらの知見を応用し、微細藻類を用いたバイオ燃料の開発や、赤潮藻類の動態(大量発生・衰退)と魚毒性に関する研究にも取り組んでいます。
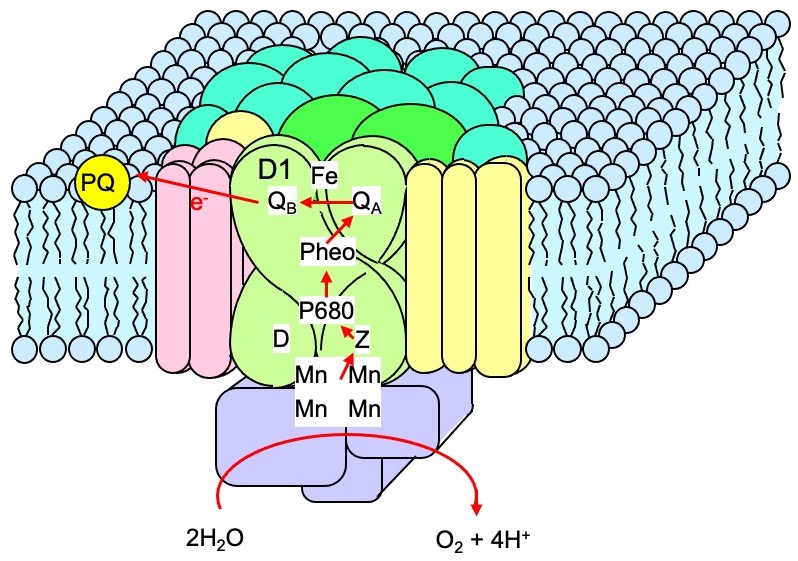
光化学系IIは光エネルギー変換を担う色素・タンパク質複合体です(図1)。しかし、光に対する感受性が高いため、強光下では容易に失活します。この現象は光化学系IIの光阻害と呼ばれ、強光下で光合成生物の生育が抑制される主な要因となります。また、強光は光化学系から大量の活性酸素を発生させ、酸化ストレスを引き起こす原因ともなります。
近年、私たちは光化学系IIの光阻害が、活性酸素によって促進されるメカニズムを明らかにしました。具体的には、活性酸素が光化学系IIの修復に必要なタンパク質の合成を翻訳過程で阻害することを発見しました。現在、私たちはシアノバクテリアなどの光合成生物を用いて、光化学系IIの光阻害とその修復の分子機構について研究を進めています。
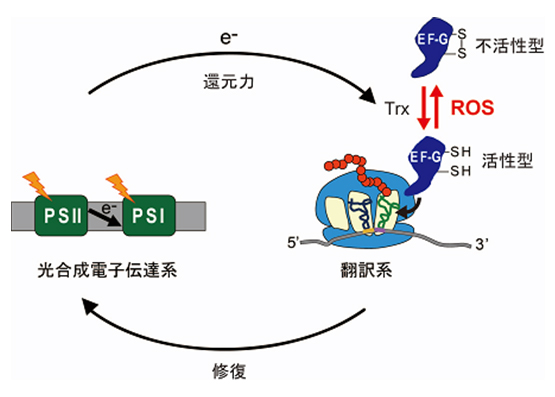
光合成における光阻害の研究を通じて、酸化ストレス下では翻訳伸長因子であるEF-GやEF-Tuが酸化されて失活し、タンパク質合成が阻害されることを明らかにしました。さらに、これらの翻訳伸長因子がチオレドキシンによって再活性化することも発見し、タンパク質合成が酸化還元(レドックス)制御を受けることを世界で初めて示しました(図2)。現在、シアノバクテリアやシロイヌナズナをモデル生物として用い、タンパク質合成系のレドックス制御における分子機構と生理学的意義の解明に取り組んでいます。
持続可能な社会を実現するためには、化石燃料に代わる再生可能エネルギーの開発が不可欠です。バイオマスエネルギーとして、微細藻類を用いたバイオ燃料の生産が試みられていますが、この生産方法における課題は、光合成効率の低下や光阻害です。代謝工学によってバイオ燃料生産能力を付与した微細藻類が開発されているものの、多くの場合、太陽光の下で培養すると、強光によって生産効率が急激に低下してしまいます。
私たちは、これまでの光合成研究で培った光防御技術をバイオ燃料生産株に応用し、強光下におけるバイオ燃料生産の高効率化と安定化を目指して研究開発を進めています。具体的には、シアノバクテリアを用いてバイオディーゼルの原料となる遊離脂肪酸を細胞外に生産させる技術(図7;下記ニュース参照)や、微細藻類ナンノクロロプシスで油脂を高生産させる技術(図8)の開発に取り組んでいます。
赤潮はプランクトンが海洋で大量に発生する現象であり(図6)、ラフィド藻や渦鞭毛藻などの有害藻類は九州内海や瀬戸内海を中心に養殖業へ甚大な被害をもたらしています。しかし、これらの藻類の光合成を含む生理学的機能については、未解明な点が多く残されています。
私たちはこれまでの研究で、有害種の渦鞭毛藻 Karenia mikimotoi における光合成の光阻害と赤潮の衰退が密接に関連していることを明らかにしました。また、別の有害種であるラフィド藻 Chattonella 属では、魚毒性の原因物質と考えられている活性酸素の産生が、光合成や栄養欠乏によって制御されていることを発見しました。
現在、私たちは赤潮藻類における光合成の分子機構や、その大量発生・衰退、そして活性酸素発生のメカニズムを研究しています。これらの研究は、赤潮の発生予測や被害防止技術の開発に貢献すると期待されます。

光合成生物は強光環境に順化することで、光合成の強光耐性を高めます。私たちは、この強光順化に伴い、光化学系IIの修復能力が増大するとともに、特殊なカロテノイドが蓄積することを発見しました(図3)。
強光環境への順化に伴い修復能力が増大する背景には、カロテノイドの蓄積や翻訳因子の誘導によってタンパク質合成が保護されるというメカニズムが関与していることが明らかになりました。現在、私たちはシアノバクテリアやシロイヌナズナをモデル生物として、光合成の強光順化機構について研究を進めています。

光化学系II複合体は高温に弱いため、その熱失活が光合成生物の高温傷害の主な要因となります。しかし、光合成生物は外部環境の温度上昇に応答し、光化学系IIの高温耐性を増大させる能力を備えています。
近年、光化学系IIの高温耐性の増大には、特定のタンパク質や脂質の新規合成が重要な役割を担っていることが明らかになりました。現在、私たちはこれらの物質の機能解明を目指して研究を進めています。この研究成果を応用し、光合成の高温耐性を増大させることも目標としています。
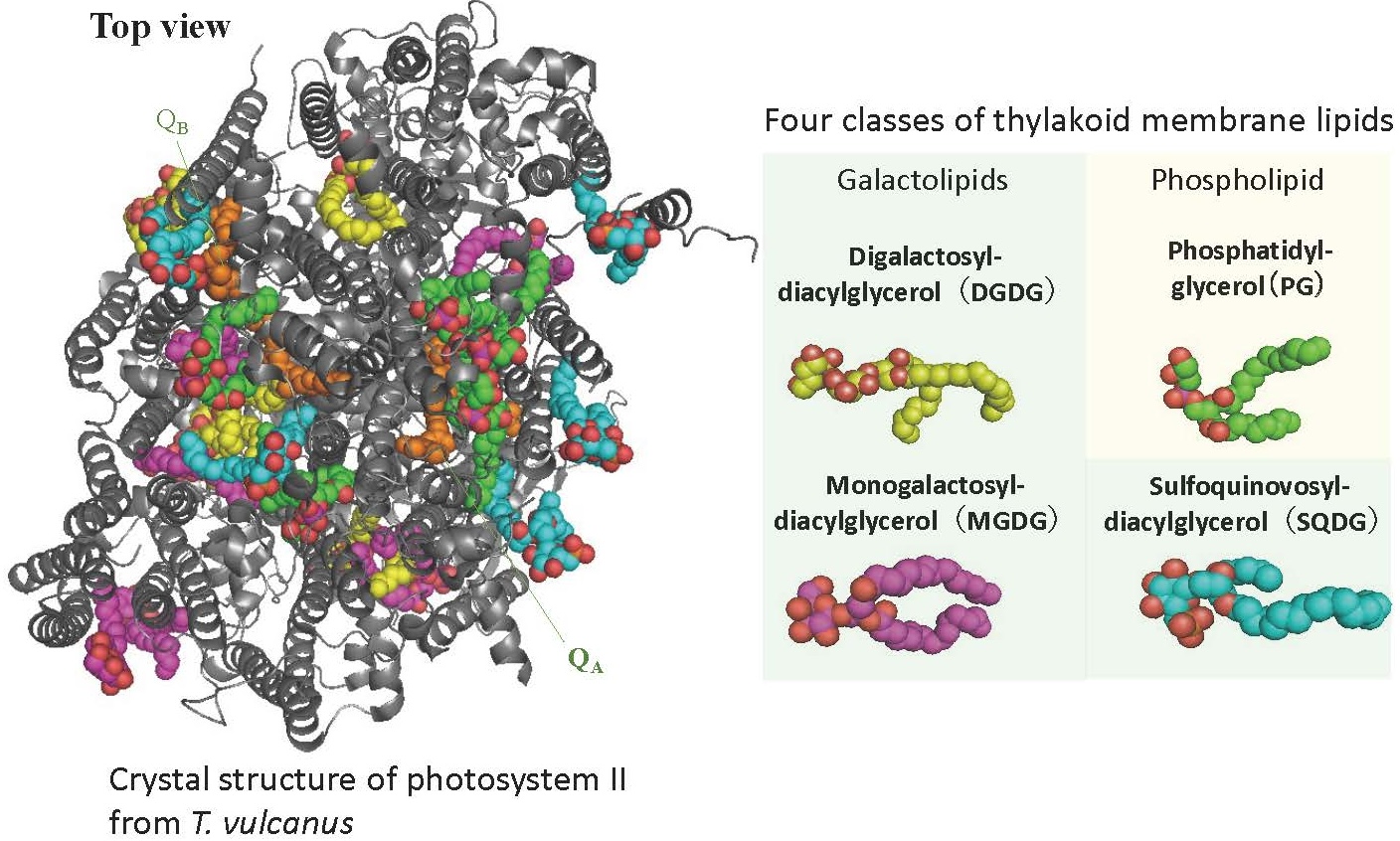
光合成が行われる場であるチラコイド膜は、4種類のグリセロ脂質から構成されています(図4)。
これらの膜脂質は、特に環境ストレス下で高速な代謝回転を示します。最近の研究では、リパーゼによって膜脂質から切り離された脂肪酸が、環境ストレスで損傷した光化学系IIの修復に重要であることが明らかになりました。現在、私たちはリパーゼ遺伝子を欠損させたシアノバクテリアや植物の変異株、あるいは化学修飾した人工脂質分子を細胞に取り込ませるケミカルバイオロジー的な手法を用いて、膜脂質の代謝回転が光合成にどのように関与しているのかを研究しています。
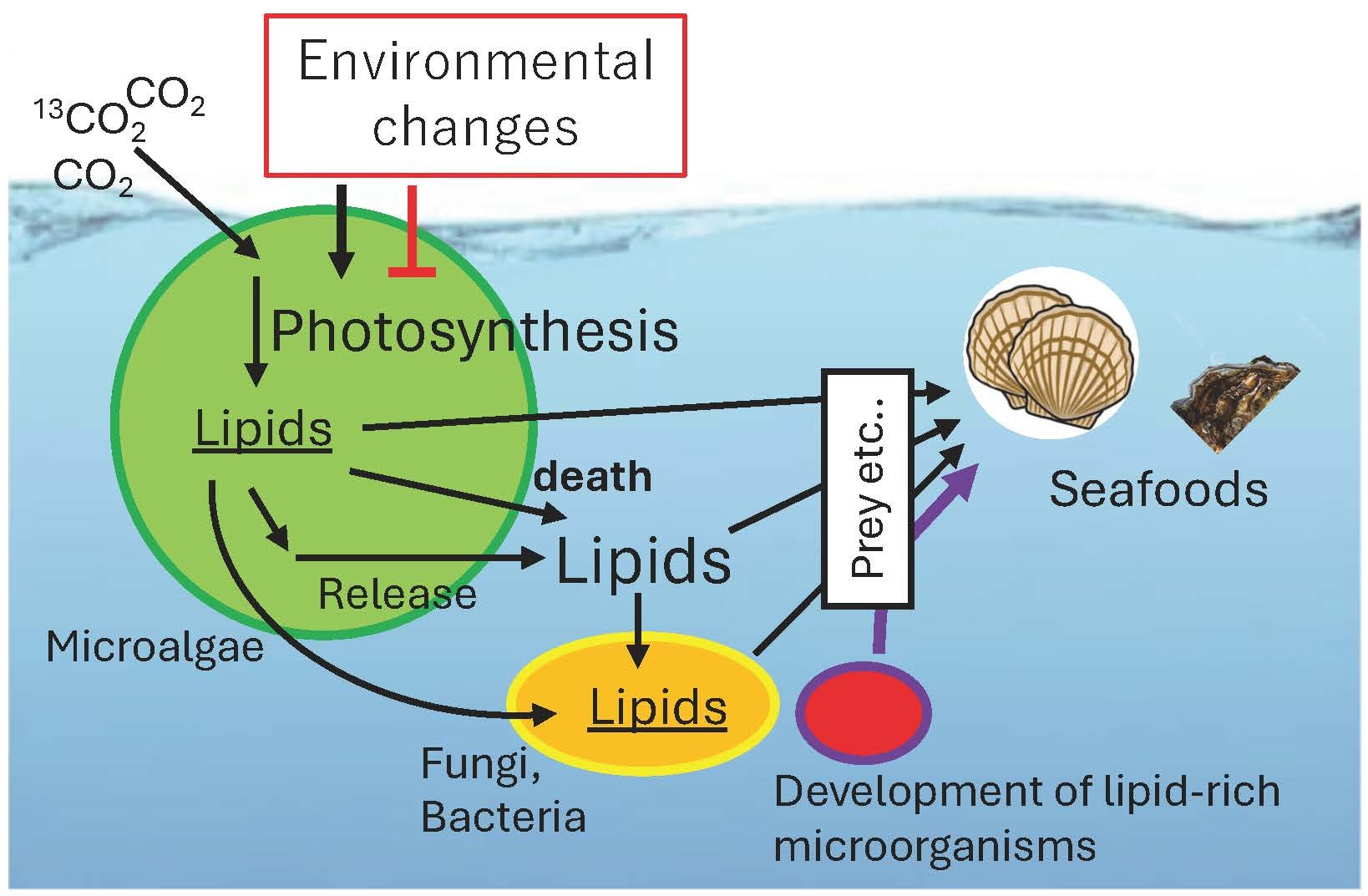
地球上の光合成活性の大部分は、緑藻、珪藻、シアノバクテリアなどの微細藻類が担っています。これらの微細藻類に含まれる脂肪酸は、動物プランクトンに捕食されたり、細胞が死滅・破裂したりすることで、生態系の中を移動します(図5)。
特に多価不飽和脂肪酸は、微細藻類によって合成されるものの、その多くは動物の生存に不可欠な成分です。これらの脂肪酸が、微細藻類からどのような経路で、どのような動物へと移行しているのかについては、未だに不明な点が多く残されています。
微生物生態系における脂質の代謝動態を解明することは、養殖魚介類の品質向上や、特定の脂質を必要とする新規微生物の単離に繋がると考え、現在、研究に取り組んでいます。
COLUMN
ミニコラム
光合成のダイナミクス
植物の葉を見ていても、短い時間なら特に変化は見られない。緑の葉っぱは緑のままだ。しかし、直射日光が当たっていると、葉っぱの中では大きな変化が起こっている。光合成装置が絶え間なく壊され復元されているのだ。損傷と修復という営みの中に生命のダイナミクスが垣間見られる。
生命を支える光合成
光合成は、太陽の光エネルギーを使って、二酸化炭素と水から有機物(糖)と酸素を生成する反応である。植物など光合成生物の生育・生存を支える最も根幹的な機能と言ってよい。生じる酸素は私たち人類を含め、好気生物の呼吸を支えているし、糖はありとあらゆる生物の炭素源となっている。そういう意味で、光合成は地球上の生命を支える営みと言って過言ではない。
光合成の損傷と修復
太陽の光エネルギーは、まず光化学系IIという色素・タンパク質複合体で化学エネルギーへと変換される。この反応に伴って水から電子が奪われ、酸素が放出される。この複合体は植物では葉緑体のチラコイド膜に存在する。光化学系IIは、その役割とは裏腹に光に対して非常に弱い。真夏の太陽光の下では30分で半分ぐらいが壊れる。しかし光合成生物は、損傷を受けた光化学系IIを速やかに修復して復元する能力をもっている。光合成機能を維持するためだ。
修復の舞台裏
光化学系IIの修復は、損傷を部分的に補修するのではなく、損傷を受けたタンパク質を丸ごと入れ替えるという大胆な戦略をとっている。損傷を受けるのは、この複合体で中心的な働きを担うD1タンパク質だが、このタンパク質を粉々に壊して取り除き、新たなD1タンパク質を遺伝子の転写・翻訳を経て一から作り直し、複合体に挿入している。たくさんのエネルギーを費やして壊れた光化学系IIを修復している。
新たな発見
私たちは、この修復プロセスが様々な環境ストレスによって阻害されることを発見した。例えば、強光の下で光合成が働けば活性酸素が大量に発生するが、活性酸素は修復のプロセスを阻害することがわかった。これは光合成の強光阻害に関する定説を覆す発見であった。さらに修復の阻害機構を生化学・分子生物学の手法を使って追求すると、タンパク質を合成する過程が活性酸素によって阻害されることがわかった。現在までに、活性酸素の標的として、EF-GやEF-Tuというタンパク質合成系の構成因子を同定している。
生命流転と展望
絶え間なく壊しては復元する−−−−。これが生物のありようなのだろう。私たちのからだも一年も経てばすべての原子が置き換わる。活性酸素や低温など環境ストレスは、修復(復元)の方を抑える働きがある。このストレス作用が光合成に特異的なのかどうか今後の課題だ。また、修復能力を強化して光合成のストレス耐性を向上させるのも目標の一つである。
(西山佳孝 埼玉新聞『サイ・テクこらむ 知と技の発信』2016年10月5日付より)
赤潮に挑む
赤潮が発生、養殖のブリやタイが何万匹も死滅、被害額は数億円? 埼玉県にいると遠い世界の出来事のようだが、養殖業の盛んな九州や四国の内海沿岸域では、毎年夏になると赤潮の脅威にさらされる。赤潮は、植物プランクトンが大量発生して海が褐色に染まる現象で、有害種による赤潮は魚介類やノリ類の養殖に甚大な被害をもたらす。
遭遇
長年研究をしていると、たまに予期せぬ出来事に遭遇する。光合成の基礎研究を行なっている筆者が赤潮に遭遇したのもそんな出来事であった。
たまたま人の紹介で赤潮を研究している若手研究者と出会った。相手は水産分野の方で、私とは畑違いであったが、話から赤潮研究にかける情熱が伝わってきた。赤潮研究は半世紀以上前から行われているが、どのように赤潮が発生して魚を殺すか、その仕組みはほとんどわかっていないという。ぜひとも解明したいが、そのためには新しいアプローチが必要だと。赤潮の原因は植物プランクトン(すなわち光合成微生物)なので、私も何か役に立てるかもしれないと思い、意気投合した。
光合成と赤潮発生・衰退
有害赤潮を形成する植物プランクトンの中で、日本で猛威を振るっているのがラフィド藻シャットネラ属や渦鞭毛藻(うずべんもうそう)カレニア・ミキモトイなどの微細藻類だ。これらの有害藻類を研究室で培養してみた。注意深く観察してみると、これらの藻類の光合成特性と赤潮形成条件に相関があることに気が付いた。シャットネラ属は、太陽の日差しが強く、海水温が高い真夏でも海面付近で赤潮を形成する。この藻類の光合成は強光・高温に強かった。一方、カレニア・ミキモトイは、真夏の晴天下では海面下5〜10メートルでとどまり、曇天になると海面付近に現れる。この藻類の光合成は強光・高温に弱かった。
また、カレニア・ミキモトイは栄養塩不足になると、遊泳運動が止まって水面付近に定位してしまう。そこに強力な光が当たれば、光合成が光阻害を受け、細胞死につながることが分かった。実際、フィールド調査してみると、貧栄養の海域で晴天時に海面付近に浮いているカレニア・ミキモトイが光阻害を受けていたことを確認できた。赤潮衰退の一因だと考えられる。
魚毒性因子
これらの有害藻類はどのように魚を殺すのか。有害藻類が魚のえらに付着して、魚を窒息死させることが推定されているが、詳細なメカニズムはほとんど分かっていない。シャットネラ属によって死滅した魚のえらを見ると、ボロボロに損傷している。単なる窒息ではなく、まるで何かの毒物にさらされたように見える。シャットネラ属は、活性酸素の一種であるスーパーオキシドを細胞外に大量に放出する。近年、スーパーオキシド産生と魚毒性に高い相関があることが分かってきた。筆者らは、スーパーオキシド産生が光合成電子伝達に依存していることを突き止めた。また、栄養塩不足によってもスーパーオキシド産生が増大することも見出した。魚毒性メカニズムの一端が見えてきた。
SDGs
海洋資源の保全や食料の安定供給は、持続可能な開発目標(SDGs)に掲げられており、赤潮発生の予測や魚毒性メカニズムの解明が実現できれば、赤潮被害を軽減させることが可能となる。本稿を執筆している2021年夏には、九州・八代海でシャットネラ属の赤潮が発生して、シマアジの養殖が9100万円を上回る被害を受けた。基礎研究のみならず、社会の要請に応える研究も進めていきたい。
(西山佳孝 埼玉新聞『サイ・テクこらむ 知と技の発信』2021年9月1日付より)